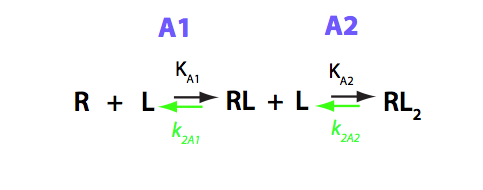
B-macro model: Two L binding to one R molecule
Evgenii Kovrigin, 12/26/2015
Derivation of differential equations describing evolution of spin concentrations
Macroscopic representation (two single-bound species are indistinguishable)
NOTE 1: This is a standard macroscopic model for the receptor binding two ligands to a receptor with two identical sites. I will work out kinetic matrices for both normal labeling scheme (R- NMR active) and try to work with reverse labeling (L- NMR active) as far as I can (EK: did not work out. Results in non-linear equations system).
NOTE 2: The mAcroscopic consideration implicitly adds together concentrations of two single-bound species. This model only applies when both single-bound species have the same chemical shift!
NOTE 3: To utilizing this model together wih microscopic models, which explicitly use concentrations of individual single-bound species, you will need to define a separate model function that formally takes microscopic constants but makes macroscopic ones for input into the B-macro formalism. For statistical effects on thermodynamic and kinetics see
IDAP/Mathematical_models/Equilibrium_thermodynamic_models/B/B_model_derivation.html
1. Reaction rates and partial conversion rates
4. Final result for the case when NMR-active nucleus is in R
6. Final result for the reverse labeling: NMR active nucleus is in L
clean up workspace
reset()
We distinguish reaction rates ( Rate, elementary reaction acts per unit time) and conversion rates (dc/dt, number of moles of specific species consumed/produced per unit time). Conversion rates, dc/dt, for species are related to reaction rates, Rate, through molecularity coefficients.
We also distinguish here partial conversion rates from net (overall) conversion rates. The net conversion rate is actual rate of change in measured concentration of the species. Partial conversion rate is the conversion rate of the species along a specific path of the reaction mechanism. By summing all partial conversion rates of the species one obtains the net conversion rate of that species.
Definitions
Equilibrium concentration of R
Req;
assumeAlso(Req > 0); assumeAlso(Req, R_);
![]()
Equilibrium concentration of ALL single-bound species, RL
RLeq;
assumeAlso(RLeq > 0); assumeAlso(RLeq, R_);
![]()
Equilibrium concentration of RL2
RL2eq;
assumeAlso(RL2eq > 0); assumeAlso(RL2eq, R_);
![]()
Equilibrium ligand concentration
Leq;
assumeAlso(Leq > 0); assumeAlso(Leq, R_);
![]()
Transition A1 :
R + L = RL
Forward macroscopic rate constant:
k_1_A_1;
assumeAlso(k_1_A_1 >0);
assumeAlso(k_1_A_1, R_);
![]()
Reverse macroscopic rate constant:
k_2_A_1;
assumeAlso(k_1_A_1 >0);
assumeAlso(k_1_A_1, R_);
![]()
Transition A2:
R + RL = RL2
Macroscopic constants
k_1_A_2;
assumeAlso(k_1_A_2 >0);
assumeAlso(k_1_A_2, R_);
![]()
Reverse rate constant
k_2_A_2;
assumeAlso(k_1_A_2 >0);
assumeAlso(k_1_A_2, R_);
![]()
Write reaction rates for the properly balanced reactions equations:
Transition A1
1. R + L => LR
a reaction rate
eq1_1a:= Rate_1_A_1 = k_1_A_1*Req*Leq
![]()
The partial conversion rate of R (one reaction act USES one molecule of R)
eq1_1b:= dcRdt_1_A_1 = Rate_1_A_1 * (-1)
![]()
the final form
eq1_1c:=eq1_1b | eq1_1a
![]()
The partial conversion rate of RL (one reaction act MAKES one molecule of RL)
eq1_1d:= dcRLdt_1_A_1 = Rate_1_A_1 * (1)
![]()
the final form
eq1_1e:=eq1_1d | eq1_1a
![]()
The partial conversion rate of L (one reaction act USES one molecule of L)
eq1_1f:=dcLdt_1_A_1 = Rate_1_A_1 * (-1)
![]()
the final form
eq1_1g:= eq1_1f | eq1_1a
![]()
2. Ligand dissociation from RL
R + L <= RL
a reaction rate
eq1_2a:= Rate_2_A_1 = k_2_A_1 * RLeq
![]()
The partial conversion rate of R (one reaction act MAKES one molecule of R)
eq1_2b:= dcRdt_2_A_1 = Rate_2_A_1 * (1)
![]()
the final form
eq1_2c:= eq1_2b | eq1_2a
![]()
The partial conversion rate of RL (one reaction act USES one molecule of LaR)
eq1_2d:= dcRLdt_2_A_1 = Rate_2_A_1 * (-1)
![]()
the final form
eq1_2e:= eq1_2d | eq1_2a
![]()
The partial conversion rate of L (one reaction act MAKES one molecule of L)
eq1_2f:= dcLdt_2_A_1 = Rate_2_A_1 * (1)
![]()
the final form
eq1_2g:= eq1_2f | eq1_2a
![]()
Transition A2
3. Ligand binding at the second site
RL + L => RL2
a reaction rate
eq1_5a:= Rate_1_A_2 = k_1_A_2 * RLeq * Leq
![]()
The partial conversion rate of RL (one reaction act USES one molecule of RL)
eq1_5b:= dcRLdt_1_A_2 = Rate_1_A_2 * (-1)
![]()
the final form
eq1_5c:= eq1_5b | eq1_5a
![]()
The partial conversion rate of RL2 (one reaction act MAKES one molecule of RL2)
eq1_5d:= dcRL2dt_1_A_2 = Rate_1_A_2 * (1)
![]()
the final form
eq1_5e:= eq1_5d | eq1_5a
![]()
The partial conversion rate of L (one reaction act USES one molecule of L)
eq1_5f:= dcLdt_1_A_2 = Rate_1_A_2 * (-1)
![]()
the final form
eq1_5g:= eq1_5f | eq1_5a
![]()
4. Ligand dissociation from RL2
RL + L <= RL2
a reaction rate
eq1_6a:= Rate_2_A_2 = k_2_A_2 * RL2eq
![]()
The partial conversion rate of RL2 (one reaction act USES one molecule of RL2)
eq1_6b:= dcRL2dt_2_A_2 = Rate_2_A_2 * (-1)
![]()
the final form
eq1_6c:= eq1_6b | eq1_6a
![]()
The partial conversion rate of RL (one reaction act MAKES one molecule of RL)
eq1_6d:= dcRLdt_2_A_2 = Rate_2_A_2 * (1)
![]()
the final form
eq1_6e:= eq1_6d | eq1_6a
![]()
The partial conversion rate of L (one reaction act MAKES one molecule of L)
eq1_6f:= dcLdt_2_A_2 = Rate_2_A_2 * (1)
![]()
the final form
eq1_6g:= eq1_6f | eq1_6a
![]()
To define evolution of the species we need to compute concentrations as a function of time. To this end, we will write differential equations for conversion rates of all species.
In a reversible process both forward and reverse reaction occur simultaneously. Thus, the net conversion rate of the species is a sum of partial conversion rates resulting from forward and reverse reactions along all branches.
Summarize conversion rates and check them against the reaction schematic
A1 transition
eq1_1c; eq1_1e; eq1_1g
![]()
![]()
![]()
eq1_2c; eq1_2e; eq1_2g
![]()
![]()
![]()
A2 transition
eq1_5c; eq1_5e; eq1_5g
![]()
![]()
![]()
eq1_6c; eq1_6e; eq1_6g
![]()
![]()
![]()
=> All seem correct
Net conversion rates
(sum all conversion rates looking at the reaction scheme)
R
eq3_1a:= dcRdt_N = dcRdt_1_A_1 + dcRdt_2_A_1
![]()
RL
eq3_1b:= dcRLdt_N = dcRLdt_1_A_1 + dcRLdt_2_A_1 + dcRLdt_1_A_2 + dcRLdt_2_A_2
![]()
RL2
eq3_1c:= dcRL2dt_N = dcRL2dt_1_A_2 + dcRL2dt_2_A_2
![]()
L
eq3_1d:= dcLdt_N = dcLdt_1_A_1 + dcLdt_2_A_1 + dcLdt_1_A_2 + dcLdt_2_A_2
![]()
Perform substitutions to obtain net conversion rates expressed in rate constants and concentrations
R
eq3_1a
![]()
list equations for corresponding rates
eq1_1c;eq1_2c;
![]()
![]()
substitute
eq3_2a:= eq3_1a | eq1_1c | eq1_2c ;
![]()
RL (both single-bound species concentrations a lumped together!)
eq3_1b
![]()
list equations for corresponding rates
eq1_1e;eq1_2e;eq1_5c;eq1_6e;
![]()
![]()
![]()
![]()
substitute
eq3_2b:= eq3_1b | eq1_1e | eq1_2e | eq1_5c | eq1_6e;
![]()
RL2
eq3_1c
![]()
list equations for corresponding rates
eq1_5e;eq1_6c;
![]()
![]()
substitute
eq3_2c:= eq3_1c | eq1_5e | eq1_6c ;
![]()
L
eq3_1d
![]()
list equations for corresponding rates
eq1_1g;eq1_2g;eq1_5g;eq1_6g;
![]()
![]()
![]()
![]()
substitute
eq3_2d:= eq3_1d | eq1_1g | eq1_2g | eq1_5g | eq1_6g;
![]()
Summarize the derivation results
eq3_2a;eq3_2b;eq3_2c
![]()
![]()
![]()
Assign order to species
eq5_1a:= Req = C1;
eq5_1b:= RLeq = C2;
eq5_1c:= RL2eq = C3;
![]()
![]()
![]()
Assign the same order to the net rates
eq5_2a:= dcRdt_N = dC1dt;
eq5_2b:= dcRLdt_N = dC2dt;
eq5_2c:= dcRL2dt_N = dC3dt;
![]()
![]()
![]()
Restate equations in terms of numbered species
eq5_3a:= eq3_2a | eq5_2a | eq5_2b | eq5_2c | eq5_1a | eq5_1b | eq5_1c
![]()
eq5_3b:= eq3_2b | eq5_2a | eq5_2b | eq5_2c | eq5_1a | eq5_1b | eq5_1c
![]()
eq5_3c:= eq3_2c | eq5_2a | eq5_2b | eq5_2c | eq5_1a | eq5_1b | eq5_1c
![]()
Prepare results for transfer to MATLAB
To avoid typing errors when transfering derived K matrix to MATLAB we type it in here and then directly test against the derivation result from above. After that the K matrix may be transfered to MATLAB by cut-and-paste of the MuPad output.
Enter the K-matrix looking at the above results (collect terms at correspondingly numbered species).
Simple rules that allow catching mistakes in K matrix derivation:
(1) a sum of each column should be zero (so each constant must appear with both positive and negative sign), and
(2) each row has to have complete pairs of constants (i.e., if k12
appears there must be k21 in the same row with an opposite sign and so on).
K_R_NMRactive:=matrix(3,3,[
[ -Leq*k_1_A_1, k_2_A_1, 0 ],
[ Leq*k_1_A_1, -k_2_A_1 - Leq*k_1_A_2, k_2_A_2 ],
[ 0, Leq*k_1_A_2, -k_2_A_2 ]
]);
![matrix([[-Leq*k_1_A_1, k_2_A_1, 0], [Leq*k_1_A_1, - k_2_A_1 - Leq*k_1_A_2, k_2_A_2], [0, Leq*k_1_A_2, -k_2_A_2]])](B_macro_images/math84.png)
Create a column vector containing concentrations of species in numbered notation
P:=matrix(3,1,[C1,C2,C3])
![matrix([[C1], [C2], [C3]])](B_macro_images/math85.png)
Check correctness of the entered K matrix by multiplying with P and comparing to the above equations:
Multiply K and P:
dCdt_manual_input:= K_R_NMRactive*P
![matrix([[C2*k_2_A_1 - C1*Leq*k_1_A_1], [C3*k_2_A_2 - C2*(k_2_A_1 + Leq*k_1_A_2) + C1*Leq*k_1_A_1], [C2*Leq*k_1_A_2 - C3*k_2_A_2]])](B_macro_images/math86.png)
Collect right-hand-side parts of equations (expressions in numbered species)
dCdt_mupad:=matrix(3,1,[ rhs(eq5_3a), rhs(eq5_3b), rhs(eq5_3c)])
![matrix([[C2*k_2_A_1 - C1*Leq*k_1_A_1], [C3*k_2_A_2 - C2*k_2_A_1 + C1*Leq*k_1_A_1 - C2*Leq*k_1_A_2], [C2*Leq*k_1_A_2 - C3*k_2_A_2]])](B_macro_images/math87.png)
Compare the derivation result to manual input
dCdt_mupad=dCdt_manual_input:
normal(%);
bool(%)
![matrix([[C2*k_2_A_1 - C1*Leq*k_1_A_1], [C3*k_2_A_2 - C2*k_2_A_1 + C1*Leq*k_1_A_1 - C2*Leq*k_1_A_2], [C2*Leq*k_1_A_2 - C3*k_2_A_2]]) = matrix([[C2*k_2_A_1 - C1*Leq*k_1_A_1], [C3*k_2_A_2 - C2*k_2_A_1 + C1*Leq*k_1_A_1 - C2*Leq*k_1_A_2], [C2*Leq*k_1_A_2 - C3*k_2_A_2]])](B_macro_images/math88.png)
![]()
=> If TRUE ---the typed K-matrix is correct.
Use this K-matrix (copy-paste output to MATLAB)
K;
![]()
The Bloch-McConnell equations describe evolution of bulk magnetization of a sample, which is proportional to the number of spins found in every specific magnetic environment. In case when NMR active nucleus is in L and in the assumption of identical binding sites in this B-macro model, the RL2 contains two spins in the identical environment, therefore the amount of magnetization from spins in the environment of RL2 is proportional to the doubled equilibrium concentration of a dimer.
Summarize the derivation results for L species concentrations
eq3_2b;eq3_2c;eq3_2d;
![]()
![]()
![]()
Assign order to species and replace concentrations of molecules with concentrations of spins in them
eq6_1a:= Leq = C1;
eq6_1b:= RLeq = C2;
eq6_1c:= RL2eq = C3/2;
![]()
![]()
![]()
Assign the same order to the net rates and replace concentrations of molecules with concentrations of spins in them
eq6_2a:= dcLdt_N = dC1dt;
eq6_2b:= dcRLdt_N = dC2dt;
eq6_2c:= dcRL2dt_N = dC3dt/2;
![]()
![]()

Restate equations in terms of numbered species
eq3_2d;
eq6_3a:= eq3_2d | eq6_2a | eq6_2b | eq6_2c | eq6_1a | eq6_1b | eq6_1c
![]()

eq3_2b;
eq6_3b:= eq3_2b | eq6_2a | eq6_2b | eq6_2c | eq6_1a | eq6_1b | eq6_1c
![]()

eq3_2c;
eq6_3c:= eq3_2c | eq6_2a | eq6_2b | eq6_2c | eq6_1a | eq6_1b | eq6_1c
![]()
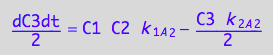
End of story for now. This is not a system of linear equations. It cannot be represented as multiplication of the K matrix and column of populations! To use such a system we need to re-derive the Bloch-McConnell equations using more sophisticated mathematics. Leave it for future...
I derived differential equations governing spin populations in the "normal" labeling scheme---when NMR-active spin is in the R. The K matrix has been prepared for transferring to MATLAB. The "reverse" labeling scheme produces non-linear system of equations---incompatible with the standard Bloch-McConnell equations and its solution using matrix algebra.